
Evolution of cooperative resistance in microbial populations subject to fluctuating environments
Why does cooperation abound in Nature? Why are there so many coexisting species? These are issues of paramount importance and challenges to Darwinian evolution [1]. In many microbial communities, it has been found that cooperative behaviour is associated with environmental changes, and with the interdependence of their size and composition (eco-evolutionary dynamics) [2].
Microbial communities evolve in volatile environments that often fluctuate between mild and harsh conditions, e.g. with sudden and radical changes in the concentration of toxins and nutrients. The resulting environmental variability greatly influences the population evolution, and the ability of species to cooperate and coexist. These are key characteristics of ecosystems that have direct applications in subjects of great societal concern, like the evolution of antimicrobial resistance (AMR) [3]. Often the mechanism of drug resistance can be viewed as a cooperative behaviour and is critically influenced by environmental and demographic fluctuations [4]. In fact, when treatments reduce a microbial community to a very small size, but fail to eradicate the microorganisms resistant to the drugs, resistant cells may replicate and restore infection, hence leading to the spread of antibiotic resistance. Moreover, within a small population, chance fluctuations may also lead to the extinction of resistant cells. In this context, central questions in the effort of modelling the evolution of AMR are: How does the joint effect of environmental variability and demographic fluctuations affect the coexistence of cells resistant and sensitive to antibiotics? How does the fraction of resistant cells vary with environmental conditions and can possibly be eradicated?
Classically, population dynamics is studied using ordinary differential equations in static environments. However, evolution is often drastically affected by fluctuations and population structure. In this project, we will address the challenge of mathematically modelling the evolution of microbial communities subject to environmental and demographic fluctuations (in time and/or space). Chiefly motivated by the application to the evolution of AMR, we will focus on models consisting of species in the presence of varying toxin and nutrient concentrations. The main features of our modelling approach, based on a combination of analytical and computational techniques, are the fluctuating size of interacting populations, and the coupling of demographic and environmental fluctuations [2,5,6,7].
Objectives & methods
By combining nonlinear dynamics and evolutionary game theory with stochastic methods and extensive computer simulations [2,5,6,7], we will study the eco-evolutionary dynamics of cooperative AMR in a well-mixed and life-cycle metapopulation settings. Our approach builds on the experience gained in the current EPSRC-NSF project “Eco-Evolutionary Dynamics of Fluctuating Populations”, and in particular from the collaboration with biologists at Imperial College, London with whom our models have been developed and who are currently probing our theoretical predictions.
1. Eco-evolutionary dynamics of cooperative AMR in well-mixed settings
The mechanism of AMR can often be regarded as a cooperative behaviour that can be affected by environmental variability and fluctuations. Resistant cells may carry resistance plasmids expressing a “resistance enzyme” that is able to inactivate the drug, offering them immunity in exchange for the metabolic cost for carrying the plasmid. On the other hand, sensitive cells do not pay the metabolic cost of carrying resistance plasmids but their growth is severely hampered when they are exposed to the antibiotic. Yet, above a certain concentration threshold of resistant cells (and enzymes), the protection offered by the “resistance enzymes” is shared with sensitive bacteria that pay no metabolic cost for it, yielding a cooperative behaviour, see Figure below from Refs. [4] and [6]. Hence, below the cooperation threshold, only the resistant cells benefit from the resistance enzymes: They outcompete sensitive bacteria and spread. However, when the concentration of resistant cells exceeds the cooperative threshold, enzymes become a public good and protect also the sensitive cells that then have a fitness advantage and outgrow resistant bacteria.
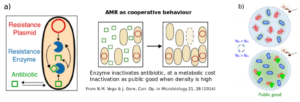
- How does the abundance of resistant cells change in time and with the environment?
- When and how can resistant cells be eradicated?
Some aspects of these problems have been considered in Refs. [6,7], and are being studied experimentally by our collaborators at Imperial College, London. Here, we will introduce new realistic modelling ingredients: (i) the concentration of antibiotics is not constant but decreases with that of resistant cells; (ii) the carrying capacity and fitness of the species both vary with the environmental conditions.
2. Metapopulation life-cycle modelling of cooperative AMR evolution
The life cycle of some microbial communities (e.g. Pseudomonas) can be described as a metapopulation model in which a large colony regularly splits into small groups of time-varying sizes. These grow independently and are reunited to form a new generation, restarting the cycle [8]. While metapopulation models have mostly been investigated in static environments, we will study the eco-evolutionary above AMR cooperative behaviour in a life-cycle metapopulation setting with time-varying concentrations of toxin and resources. We will be particularly interested in analysing the differences from, similarities with, the well-mixed case when cells are allowed to migrate from one group to another (dispersal).
References
[1] E. Pennisi, How Did Cooperative Behavior Evolve?, Science 309, 93 (2005); What Determines Species Diversity? Science 309, 90 (2005).
[2] K. Wienand, E. Frey, and M. Mobilia, Evolution of a Fluctuating Population in a Randomly Switching Environment, Phys. Rev. Lett. 119, 158301 (2017); Eco-evolutionary dynamics of a population with randomly switching carrying capacity, J. R. Soc. Interface 15, 20180343 (2018).
[3] J. O’Neill, Tackling drug-resistant infections globally: Final report and recommendations (2016).
[4] E. A. Yurtsev, H. X. Chao, M. S. Datta, T. Artemova, and J. Gore, Bacterial cheating drives the population dynamics of cooperative antibiotic resistance plasmids, Mol. Syst. Biol. 9, 683 (2013). N.M. Vega and J Gore, Collective antibiotic resistance: mechanisms and implications. Current Opinion in Microbiology, 21,28 (2014).
[5] A. Taitelbaum, R. West, M. Assaf, and M. Mobilia, Population Dynamics in a Changing Environment: Random versus Periodic Switching, Phys. Rev. Lett. 125, 048105 (2020); Evolutionary dynamics in a varying environment: Continuous versus discrete noise, Phys. Rev. Research 5, L022004 (2023).
[6] L. Hernández-Navarro, M. Asker, A. M. Rucklidge, and M. Mobilia, Coupled environmental and demographic fluctuations shape the evolution of cooperative antimicrobial resistance, bioRxiv 2023.07.06.547929 (to appear in J. R. Soc. Interface).
[7] M. Asker, L. Hernández-Navarro, A. M. Rucklidge, and M. Mobilia, Coexistence of Competing Microbial Strains under Twofold Environmental Variability and Demographic Fluctuations, arXiv:2307.06314.
[8] G. Velicer & M. Vos, Sociobiology of the Myxobacteria, Annu. Rev. Micro. 63, 599 (2009); J. S. Chuang, O. Rivoire, and S. Leibler, Simpson’s paradox in a synthetic microbial system, Science 323, 272 (2009).